A brief look at the family Leporidae (hares and rabbits)
Keywords: Leporidae, Oryctolagus, Lepus, Sylvilagus, anatomy, physiology, reproduction, uterus, ovaries, tract, vagina, rabbits, hares
A fascinating group of mammals, some familiar to veterinary and animal science, others not. Note that rabbits and hares belong to the order Lagomorpha, of which there are two families: Leporidae and Ochotonidae (pikas, mouse hares, or conies)
As mentioned in the LORI entry on semen collection in rabbits, rabbit farming is underdeveloped in North America compared with Europe, Asia and South America. Therefore rabbits are superficially addressed in most animal science and veterinary curricula in North America. However, rabbits deserve attention as important sources of meat and fiber and their unique value as subjects for scientific study (See: Foote and Carney, 2000).
Vocabulary:
Lagamorph: A member of the order Lagamorpha
Leporid: A member of the family Leporidae, comprising the rabbits and hares.
Kindling: The name given to parturition in rabbits and hares.
Kits: Perinatal rabbits and hares (the term "pup" is reserved for rats, mice and dogs).
Doe: Adult female Leporid
Buck: Adult male Leporid.
Nests and burrows: Hares (always wild) live in nests, above ground. Wild rabbits such as Cottontails live in burrows.
| Introductory notes: Rabbits and hares are all members of the family Leporidae. This family includes the domestic rabbits (Oryctolagus cuniculus), hares (many Lepus spp) and brush rabbits (many Sylvilagus spp). They share similar reproductive characteristics but as donkeys are to horses, they are also distinctively different. For example, hares and brush rabbits have 48 chromosome and precocious neonates with open eyes, able to ambulate efficiently. By contrast, rabbits have 44 chromosomes and altricial young, blind, hairless and unable to ambulate. Their eyes open at about 10 days postpartum. In the course of this entry, the author will entertain other differences between hares and rabbits. |

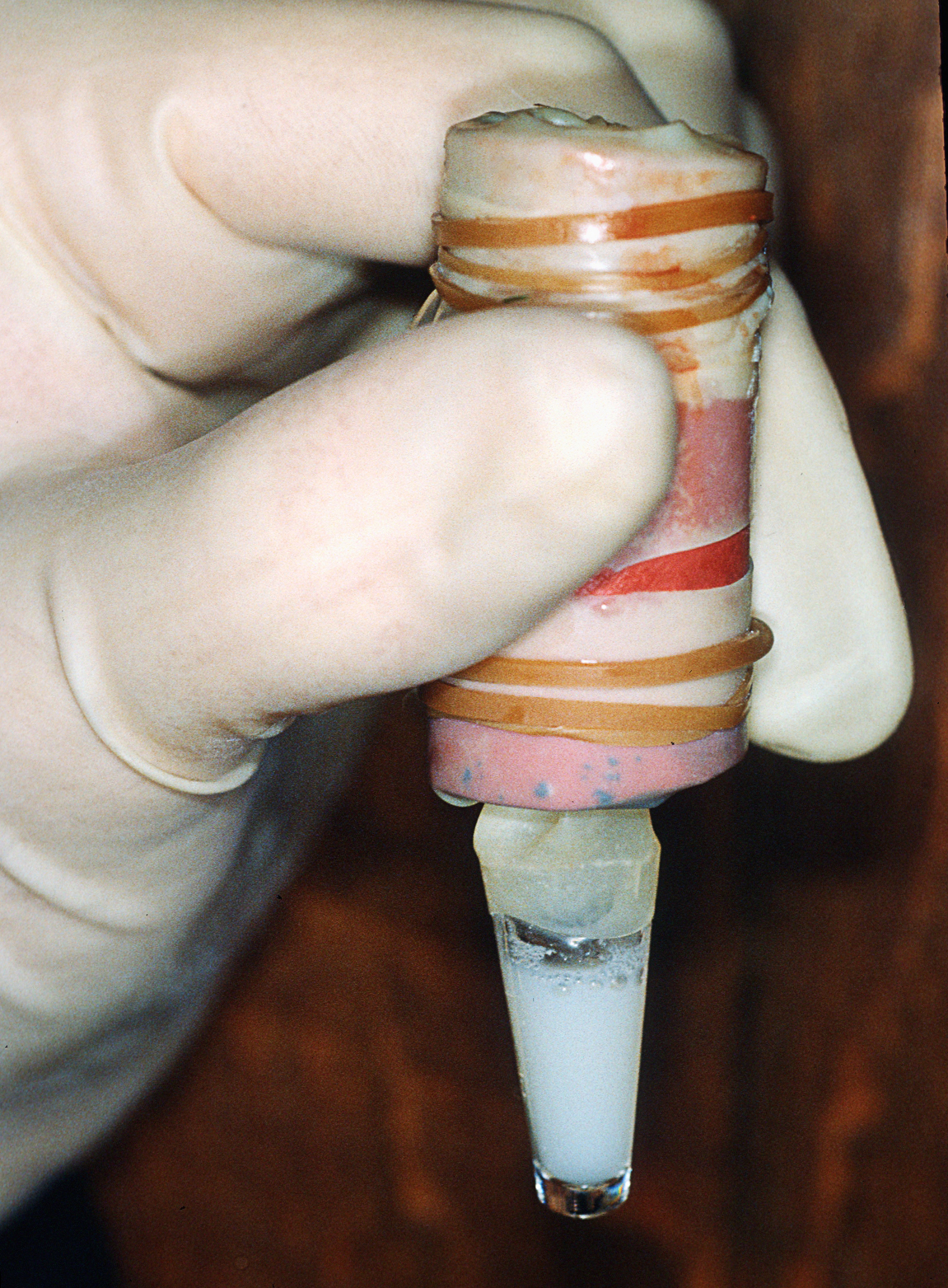
Figure 1. The reproductive tract of a Snowshoe hare (Lepus Americanus) that was approximately 14 days pregnant. The ~20 day old fetus at left is that of a domestic rabbit (Oryctolagus cuniculus). The 14 day pregnancy shows the bulbous embryo-placental units typical of rabbits and hares. Note too, that the ovaries are not covered by ovarian bursae. Corpora lutea of pregnancy are visible within the ovaries. Finally, note the double cervix (inset at right); typical for all Leporids (as well as rats and mice). The vagina (not shown) is about 10 to 12 cm long. Image size: 1344 x 831 px
| A crash course on rabbit and hare reproduction:
1. Puberty occurs between 4 and 7 months of age; later in large rabbit breeds such as the Flemish White.
2. Although commercial rabbits breed throughout the year, hares and wild rabbits (Sylvilagus spp; Cottontails) are like cats. i.e. they respond to increasing day length and breed mostly during spring and early summer.
3. Like cats, rabbits and hares do not have discrete estrous cycles. Instead, as reflected by vaginal cytology, follicle waves mature every 4 to 5 days (~ 7 days in hares).
4. Although spontaneous ovulation can occur, ovulation is generally induced by copulation (mounting in the absence of intromission is also thought to be important). Ovulation occurs about 10 hours post copulation. GnRH treatment is essential for satisfactory pregnancy rates when AI is used.
5. Pregnancy diagnosis is accurate at 10 to 14 days post-breeding using transabdominal palpation. Optimal first service conception rates should be greater than 70% i.e. 70% of does should be pregnant after a single breeding.
6. Gestation is longer in hares than rabbits; about 42 days vs. 30 days. This explains why hares have precocious young and rabbits, altricial young.
7. Parturition occurs in the morning. Anterior, posterior and even breech presentations are normal. Usually kindling is complete in less than 30 minutes but occasionally, kits (like kittens) may be born hours or even one or two days apart. Kits are usually born separated from their umbilical cords but maternal biting of cords has been described.
8. Cannibalism (infanticide) is fairly common in young rabbits, especially under stressful conditions.
9. Rabbits have an average of six to 10 kits per litter. Does have 8 to 10 mammary glands and nurse their kits once per day.
10. Pseudopregnancy occurs after sterile mating in both hares and rabbits but does not normally occur in the absence of mating as in dogs and cats. Like pregnancy itself, pseudopregnancy is slightly longer in hares than rabbits i.e. 24 days vs. 17 days. As is sometimes seen in bitches, nesting behavior will occur during pseudopregnancy. In the case of Leporids this involves pulling of abdominal hairs and gathering of straw etc.
11. Pregnancies with one or two fetuses may be significantly longer than otherwise (as is the situation with cats and dogs). One can usually diagnose retained fetuses by palpation. Cesarean section may be required in such cases.
12. Rabbits can have fertile copulation soon after birth and become pregnant in under a week after parturition (kindling ). This is possible because rabbits have active ovaries at the time of kindling and uterine involution is largely complete within 48 hours of parturition. Consider that other domestic animals have ovaries devoid of significant follicle growth at parturition.
13. Conception soon after parturition is remarkable in light of the intimate placentation in Leporids. It is hemochorial i.e. more intimate than any other domestic animal (see figure 2).
In commercial operations, does are re-bred between 2 and 6 weeks after kindling. This will provide 4 to 6 litters per year.
14. Weaning in commercial operations usually occurs at approximately 6 weeks, when the doe has already been pregnant for some time.
15. Rabbits are usually processed for meat production at 3 to 4 months of age and breeding stock rabbits are seldom kept for longer than 3 to 4 years in commercial units.
|

Figure 2. Placentation typical of a rabbit or hare; a modified amalgum from several sources. Placentation is bi-discoidal then discoidal and intimate, developing mesometrially (although anti-mesometrial in early gestation). Placentation is hemochorial (also referred to as hemobichorial) with giant cells forming in the endometrium, similar to equine eCG-producing cells. Note the large exocelom, similar to that in carnivores. Image size: 896 x 733 px
Selected references:
Benirschke. K. Comparative placentation. http://placentation.ucsd.edu/rabbits.htm
Chavatte-Palmer P. et al. 2008. In utero characterization of fetal growth by ultrasound scanning in the rabbit. Theriogenology. y 69: 859–869
Dickie, E. 2011.Dystocia in a rabbit (Oryctolagus cuniculus). Can. Vet. J. 52: 80-83.
Boumahdi, Z. et al. 20009 Behavior at birth and anatomo-histological changes studies of uteri and ovaries in the post partum phase in rabbits. European J. of Scientific Res. 34: 474-484
Foote, R.H. and Carney E.W. 2000.The rabbit as a model for reproductive and developmental toxicity studies
Foxcraft, G.R. and Hasnain, H. 1973. Effects of suckling and time to mating after parturition on reproduction in the domestic rabbit. J. Reprod. Fert. 33:367-377
International rabbit reproduction group. 2005.Guideline for the handling of rabbit bucks and semen. World rabbit.Sci. 13:71-91
Roellig, K. et al 2010. Superconception in mammalian pregnancy can be detected and increases reproductive output per breeding season. Nat Commun. 2010 Sep; 1(6): 1–7.
Torres, S. et al. 1977. Fertility factors in lactating rabbits mated 24 hours and 25 days after parturition. Ann. biol. anim. Biochem. Biophysics. 17:63-69
Weisbroth et al Eds. 2013. The Biology of the Laboratory Rabbit. ISBN 1483270319